IX. The Biosynthesis Of Complex Lipids
Description
This section is from the book "The Scientific Contributions Of The Ben May Laboratory For Cancer Research", by The University of Chicago. Also available from Amazon: The Scientific Contributions Of The Ben May Laboratory For Cancer Research.
IX. The Biosynthesis Of Complex Lipids
In 1952 there was no information on the detailed chemical pathways leading to the biosynthesis of complex lipids such as lecithin. Since compounds of this type are found in every kind of living cell, this obviously represented a major gap in our knowledge of intermediary metabolism. Accordingly, a series of investigations was undertaken to work out the enzymatic processes leading to the formation of lecithin as a representative phospholipid.
This work led to the discovery of the function of cytidine coenzymes in the biosynthesis of phospholipids. The structure of the first such compound to be discovered, cytidine diphosphate choline, is shown in Figure 31.
A detailed scheme for the biosynthesis of lecithin from simple precursors was established (Fig. 32). A similar and parallel series of reactions was found to lead to the synthesis of phosphatidylethanolamine in the living cell.
A pathway for the de novo formation of triglycerides was also shown to involve phosphatidic acids and D-a,β-diglycerides as intermediates (Fig. 32).
L-A-Glycerophosphate As An Intermediate In The Biosynthesis Of Phospholipids
In a study (122) of the incorporation of P32 into the phospholipids of a particulate enzyme preparation from liver, it was found that a preliminary conversion of P32 to ATP was necessary, and the addition of free glycerol approximately doubled the rate of incorporation, suggesting that reaction (a) of Figure 32 is an essential step. Confirmation of this hypothesis came with the direct demonstration of the conversion of synthetic a-glycero-phosphate labeled with P32 to a lipid with chromatographic properties similar to that of a phosphatidic acid. Unlabeled a-glycerophosphate also acted as an effective isotope diluent, confirming the fact that it is an obligatory intermediate in the formation of the labeled phospholipid.
Purification And Properties Of Glycerokinase
Glycerokinase, the enzyme catalyzing reaction (a) of Figure 32, was partially purified (4). The phosphorylation of glycerol, an optically inactive molecule, takes place in an asymmetrical fashion, yielding L-o-glycero-phosphate only (5). Other interesting properties of the enzyme are its heat stability in the presence of glycerol and its ability to catalyze the phosphorylation of dihydroxyacetone and of L-glyceraldehyde, as well as that of glycerol.
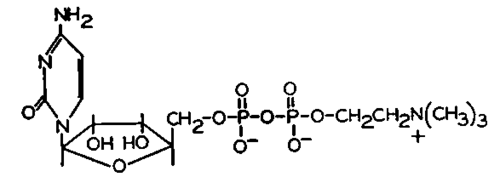
Fig. 31.-The structure of cytidine diphosphate choline.

Fig. 32.-Pathways for the enzymatic synthesis of phospholipids and triglycerides.
Direct evidence that glycerokinase mediates an important step in the conversion of glycerol to glycogen was also obtained (5).
Phosphatidic Acid Phosphatase
The important studies of Romberg and Pricer (J. Biol. Chem., 204:345, 1953) as well as the work described above had made it clear that phosphatidic acids are active intermediates in the biosynthesis of lipids. The next step in working out the pattern of reactions shown in Figure 32 was the discovery of a specific phosphatase which cleaves the phosphate moiety from a phosphatidic acid with the formation of inorganic phosphate and D-a,β-diglyceride (188). This reaction appears to be essential for the conversion of the glyceride portion of phosphatidic acids to triglycerides, as well as to phosphatidylethanolamine and lecithin.
Function Of Cytidine Coenzymes
A preliminary report by Kornberg and Pricer (J. Am. Chem. Soc., 74: 1617, 1952) had indicated that phosphorylcholine could be incorporated as a unit into a phospholipid which was identified by later workers as lecithin. It was suggested as a working hypothesis that phosphorylcholine might react with a phosphatidic acid to yield lecithin. However, no such reaction could be demonstrated in this laboratory (129). It was found instead that the conversion of phosphorylcholine to lecithin by particulate enzymes from rat liver was stimulated by amorphous ATP (Pabst lot 116) but not by crystalline ATP (Pabst lot 120). The activity of the amorphous ATP was traced to its contamination by the true cofactor, CTP. This, in turn, led to the discovery of cytidine diphosphate choline (Fig. 31), which marked the first appearance of a cytidine coenzyme in a reaction of major metabolic importance.
Evidence that the function of cytidine coenzymes in the synthesis of lecithin is as shown in Figure 32 was presented in a report (130) which also described the enzymatic synthesis of phosphatidylethanolamine by a corresponding series of reactions, involving phosphorylethanolamine and CDP-ethanolamine in the place of phosphorylcholine and CDP-choline, respectively. A detailed study of each of the reactions shown was carried out in later work, which fully corroborated the postulated schemes.
It was also possible to demonstrate the net synthesis of lecithin in vitro and to show directly the dependence of this process on the dephosphoryla-tion of phosphatidic acid (188).
Synthesis Of Cdp-Choline, Cdp-Ethanolamine And Related Compounds
The methods developed in the laboratories of Khorana and of Todd for the synthesis of diesters of pyrophosphoric acid, using N,N'-dicyclohexyl-carbodiimide as a general reagent for this purpose, were applied to the synthesis of CDP-choline, CDP-ethanolamine and related compounds (125). The availability of these pure synthetic materials in various labeled forms greatly facilitated further progress.
Enzymatic Synthesis Of Triglycerides
When it became clear that the enzymatic synthesis of lecithin proceeded through stages involving the formation of phosphatidic acids and D-a,0-diglycerides, the hypothesis was put forward that these compounds might also be important intermediates in the synthesis of triglycerides. Evidence in support of this idea was obtained (187) by the demonstration of the net synthesis of triglycerides according to reaction (h) (Fig. 32), using particulate enzymes derived from chicken liver. Some properties of the enzyme catalyzing this reaction were described in a later publication (Weiss, S. B.; Kennedy, E. P.; and Kiyasu, J. Y. J. Biol. Chem., 235:40, 1960).
Subsequent work in other laboratories has shown that the pathway discovered for triglyceride synthesis in liver also occurs in other tissues such as intestinal mucosa and fat.
Continue to:
My Books

